ARCHIVE FILE
This article was published in 1959
See the original document
INSTITUTE OF INSPECTORS OF STOCK OF N.S.W. YEAR BOOK.
Some Recent Advances in Genetics from the Point of View of Animal Breeding
J. S. F. BARKER, B.Agr.Sc., Dept. of Animal Husbandry, University of Sydney
Before considering some of the recent advances in genetics, it is desirable first to talk about animal breeding as it is known today. It is well known that genetic improvement results from the selection of those animals showing superior breeding value or genetic worth, and then using them as breeding animals. However, animal breeders have been using this selection for many years and though improvements have been made, these improvements often have been quite small or negligible. This apparent contradiction has in many cases resulted in a loss of faith in the power of animal breeding to produce any worthwhile results. It can, however, be explained quite simply.
The explanation is based on a concept that has been the backbone of animal breeding theory for just over twenty years the concept of heritability. This concept was first expounded in detail by J. L. Lush of Iowa State College in the late 1930's; though the basic theory had been laid by Sewall Wright nearly 20 years earlier. Most or all of you will be familiar with the idea of heritability, but a brief review of its meaning and significance could be helpful. Heritability refers to the average influence of the genetic factors, and is used to express the fraction of the difference between groups of parents that we may expect to recover in their offspring. The greater the fraction that is inherited, the higher the heritability. For example, if a group of parents have an average production of 200 units (where the units may be weight at a certain age in beef cattle, pounds of butterfat per cow or egg production per bird), while the average of the herd or flock from which they were selected is 160 units, then the superiority of the selected parents is 40 units. If their progeny average 176 units, then the heritability of the character is:—
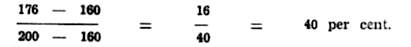
Thus, if the heritability of a given character is low, then only a small fraction of the differences between parents is recovered in their offspring. In this case, the differences between parents are due mainly to environmental factors. If the heritability were 100 per cent., all the differences between parents would be recovered in their offspring; and in this case, the breeding value of an animal coincides with its observed performance. This introduces another way of looking at heritability in that it may be considered as measuring the degree of correlation between the breeding value or genotype of an animal and its observed performance or phenotype. We generally classify the heritability estimate of a particular character into one of three classes:
(i) High — heritability greater than 30 per cent.
(ii) Medium — heritability between 15 and 30 per cent.
(iii) Low — heritability less than 15 per cent.
For characters with a high heritability, the correlation between genotype and phenotype is high enough for us to make maximum genetic progress by selecting animals on the basis of their own performance only; that is, mass or individual selection. However, when heritability is low, selection of individual performance that one is necessarily selecting those animals having the best breeding value, so that genetic improvement is consequently quite slow. This explains the contradiction mentioned earlier. When heritability is low, we need some better guide to an animal's genotype than its own phenotype and we usually get this by a consideration of the phenotypes of relatives, usually sibs (sister and/or brothers) or progeny. The average phenotypes of a sufficient number of such relatives gives a better guide to an individual's genotype than its own phenotype. Such selection is called family selection. For characters with medium heritability, we use some combination of individual and family selection.
The rate of genetic progress is determined not only by the heritability, but also by the selection differential and the generation length. The selection differential is the degree of superiority of the individuals selected for breeding over the whole population from which they came. In our previous example, the population average was 160 units and the selected parents average 200 units; so that here, the selection differential is 40 units. The generation interval is the average age of parents when the offspring are born. Progress per year is more important than progress per generation and is estimated by dividing the expected progress per generation by the generation interval.
Progress per year = Selection Differential x Heritability / Generation Interval
Care is needed when using family selection as this increases the generation interval, and the increased accuracy of selection (increased effective heritability) must more than compensate for this so that progress per year is actually increased by the use of family selection.
A knowledge of heritability and the selection differential applied allows us to predict the expected rate of genetic progress per generation. For a given selection differential each generation, we would expect this rate of progress to be linear, but that it would eventually cease when we had made the population homozygous for all desirable genes affecting the character being selected. At this stage, the population should show zero heritability. Also, there should be no tendency for the characters to regress if selection is relaxed. In all selection experiments that have been carried far enough, responses have ceased after a variable time, when a plateau is reached.
It is impossible to predict if and when a selection plateau is likely to be reached in any particular selection programme. Even when a plateau apparently has been reached. It is not possible is a temporary stage, where continued selection may result in a further improvement, or if it is a definite limit that further selection will not raise. It may be of interest, however, to look at some of the results obtained in actual selection experiments. Clayton and Robertson (1957), in selection for increased bristle number in Drosophila, in one line obtained rapid early response which ceased suddenly at generation 19, and thereafter no change. In another line, the response was slow, but continuous for 30 generations, while in the three following generations to the end of the experiment there were suggestions that response had ceased. Falconer (1955) selected for high and low body weight in mice and found the response apparently ceased at generation 22 in the high line and at generation 17 in the low line.
In many of these selection experiments, there was no or little reduction in heritability when the plateau was reached and following suspension of selection regression of the character did occur. (For example, Lerner (1954), Falconer (1955) and Clayton and Robertson (1957). Thus, in these experiments selection response ceased while genetic variation that should have been usable remained. Some possible causes of such selection plateaux are:
(i) Opposing natural selection. Natural selection against animals with extreme phenotypes results in expected selection differentials being greater than realised selection differentials. Extreme phenotypes fail to reproduce as efficiently as the individuals closer to the original mean of the population.
(ii) Reduction in heterozygosity. Certain experiments have shown at in some cases the more heterozygous individuals are fitter (or more able to survive and reproduce) than the more homozygous. As heterozygosity is reduced by selection, a stage is reached where the more homozygous individuals (the more desirable individuals) fail to reproduce as efficiently as the less homozygous ones. This idea has been developed most extensively by Lerner (1954).
In many cases, the factors responsible for the cessation of selection response are not understood and are described in general terms as physiological limits.
Most of the experiments in which selection plateaux have been observed have been done with Drosophila or mice, so that this may be thought of only theoretical interest and not of much application to animal breeding. It will become of importance in animal breeding, and probably of extreme importance, before many years. Already there are suggestions in certain livestock, notably poultry (Dickerson 1955), and it is almost certain that they will be reached in other economic animals.
The problem of major importance then will be — Is it possible to break through these selection barriers, and if so, how? The problem of overcoming natural selection against extreme phenotypes has not been solved. Certain measures to reduce the effect, viz., avoidance of inbreeding and selection at a slower than maximum rate, were suggested by Lerner (1954). Avoiding inbreeding would limit homozygosity as far as possible to the loci under selection, thus reducing the depressant effect of homozygosity per se on fitness. A moderate selection pressure would give more time for genetic and physiological adjustment as the character under selection was moved away from its original level.
Some methods for breaking through selection barriers have been suggested and are being investigated.
Mutation renews or helps to maintain genetic variation. However, spontaneous mutation is too slow to be of value in selection work, but selection within a population which has been treated in order to increase mutation rate could overcome this difficulty. Scossiroli (1954) showed that by using X-irradiation to increase mutation rate, he was able to break through a selection plateau in Drosophila and obtain further response.
Crossing of two or more unrelated strains with a similar history of selection and then selecting within the hybrid progeny may allow further progress. The different strains may have reached the same end point by different physiological means — homozygosity for different sets of genes. Falconer and King (1953) crossed two strains of mice selected for heavy body weight both of which had reached plateau. A further increase in body weight was obtained by selection within the hybrid progeny.
Another method has been suggested by the results of Waddington (1953, 1956). He showed that an invariant character (a specific wing venation character in Drosophila) could be made to vary by a suitable environmental treatment. (In this case, heat treatment of larvae). Selection on this variability built up a genotype which eventually produced the character in the absence of treatment. In this case, the environmental treatment produced the desired character. In animal breeding, considerable research will be needed to find suitable environmental treatments. It is thought by some workers, though, that it may not be necessary to use a treatment that produces desirable results in itself. Quite a lot of research is being done at the moment selecting animals under many devious environments, all quite different from the normal and all imposing stress on the animals. What these experiments will produce we have yet to see. As far as one knows, none of this has yet been published.
A method similar to that of Waddington is now under study and as far as we know, is only being studied by geneticists here in Sydney. This method, Instead of using an environmental treatment. Uses a major gene which affects the character under attack. Dun and Fraser (1958, 1959) are using this method with the mouse, and Rendel (1959) with Drosophila. Dun and Fraser have used as the invariant character "vibrissa-number", and Tabby is the major gene affecting it.
The Tabby gene was associated with a reduction of mean and increase of variance in vibrissa-number. Thus lack of normal variation was overcome by selecting within the phenotypes of heterozygous Tabby females and Tabby males. By the seventh generation of selection, the high and low lines differed by about 4 vibrissae in heterozygous Tabby females by about 3 vibrissae in Tabby males, and in normal mice by about 0.5 vibrissae. Thus, selection for changes of vibrissa number in Tabby mice has caused changes in the vibrissa number of normal mice.
Another possible method of making further improvement after a selection plateau is reached is to take advantage of heterosis by using special mating techniques, such as reciprocal recurrent select on. This method involves progeny testing and selection within a segregating population. The selection of individuals for further breeding is not based on their own phenotypes but upon the merits of their cross progenies. The method features two segregating populations, each serving as a tester for the other. Bell, Moore and Warren (1955) showed a slight superiority of this technique over simple selection, using a character in Drosophila of low heritability (viz., fecundity or number of eggs laid per female). The value of the method depends on the presence of "combining ability" for the character being selected and this may or may not be present (Comstock and Robinson, 1957).
This, then, is the general picture. We may expect to reach selection plateaux in our economic livestock. Just when this will occur in any particular case, we cannot predict. However, various methods of overcoming or breaking through these selection barriers have been suggested and are being investigated. This work is of particular significance and it is important that the theoretical knowledge of selection barriers and methods of overcoming them be developed fully before we actively strike this problem in animal breeding.
REFERENCES.
I. General References on the theory and genetic background of the concept of heritability.
LERNER, I. M. (1950) - Population Genetics and Animal Improvement. Cambridge, Cambridge University Press
LERNER, I. M. (1958)-The Genetic Basis of Selection. New York, John Wiley.
LUSH, J. L. (1945) - Animal Breeding Plans. Ames, Iowa State College Press.
II. Specific references in the Text.
BELL, A. E., MOORE, C. H. and WARREN, D.C. (1955)-The evaluation of new methods for the improvement of quantitative characteristics. Cold Spring Harbour Symp. Quant. Biol. 20: 197 - 212.
CLAYTON, G. A. and ROBERTSON, A. (1957)-An experimental check of quantitative genetical theory II. "The long term effects of selection. J. Genet. 55: 152 - 70.
COMSTOCK, R. E. and ROBINSON, H. F. (1957)-Findings relative to reciprocal recurrent selection. Proc, Int. Genet. Symp. (Tokyo and Kyoto). 1956. Tokyo, Sel. Council of Japan. pp. 461-4.
DICKERSON, G.E. (1955)-Genetic slippage in response to selection, Cold Spring Harbour Symp. Quant. Biol. 20 : 213 - 23.
DUN. R. B. and FRASER, A. S. (1958)-Selection for an invariant character - "vibrissa-number" - in the house mouse. Nature. 181 : 1018 - 9.
DUN, R. B. and FRASER, A. S. (1959)-Selection for an invariant character - "vibrissa-number" - in the house mouse. In press.
PALOONER, D. S. (1955) - Patterns of response in selection experiments with mice. Cold Springs Harbour Symp. Quant. Biol. 20: 178 - 95.
FALCONER, D. S. and KING, J. W. B. (1953)-A study of selection limits in the mouse. 1. Gener, 51 : 561 - 81.
LERNER I. M. (1954) - Genetic Homeostasis.
RENDEL J. M. (1959) - Canalisation of the acute phenotype. Evolution (in press).
SCOSSIROLI, R. E (1954)-Effectiveness of artificial selection under irradiation of plateaued populations of Drosophila melanogaster. I.U.B.S. Symp. Geneties of Population Structure, Series B No. 15 (Aurust 1954) pp. 20-23
WADDINGTON. C. H. (1953)- Genetic Assimilation of an acquired character. Evolution 7: 118 - 26.
WADDINGTON. C. H. (1956)-Genetic Assimilation of the bithorax phenotype. Evolution 10 : 1 - 13. (NOTE Paper delivered to the Annual Conference of the Institute of Veterinary Inspectors of N.S.W.: 20th March, 1959.)